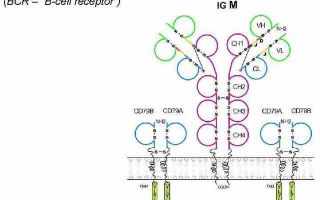


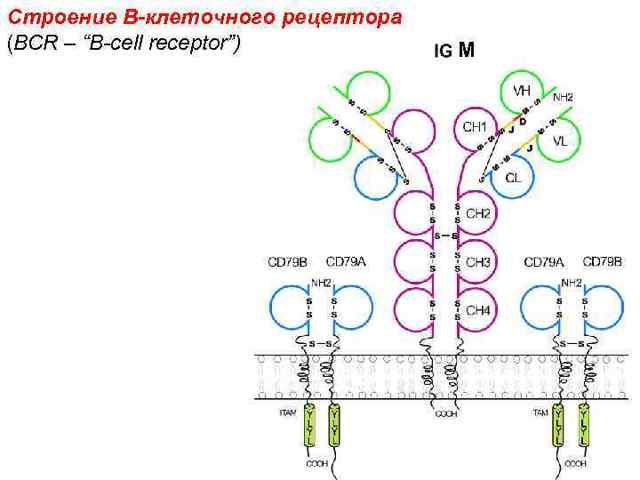
Основным процессом дифференцировки В-лимфоцитов является формирование антигенраспознающего В-клеточного рецептора!
Локализация генов иммуноглобулинов у человека и мыши
Организация зародышевых генов иммуноглобулинов
Реаранжировка (перестройка) генов D 1 -D 27
Для перегруппировки генов H- и L-цепей требуются ферменты рекомбиназы RAG-1 и RAG-2 (recombinationactivating genes).
Комбинаторное разнообразие иммуноглобулинов
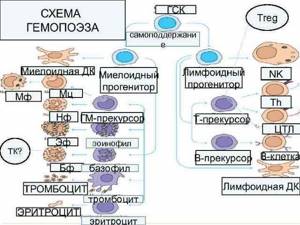
Этапы дифференцировки B-лимфоцитов
Пре В-клеточный рецептор
Отрицательная селекция В-лимфоцитов в костном мозгу: реакции незрелых клеток на распознавание аутоантигена – индукция анергии, редактирование V-генов и апоптоз
Антиген-зависимый этап дифференцировки В-лимфоцитов (гуморальный иммунный ответ)
Дифференцировка В-лимфоцита в плазматическую клетку Плазматическая клетка B-лимфоцит Утрата экспрессии генов В-клетки Блокада клеточного цикла после плазмобласта Увеличение размера, расширение цитоплазмы, развитие шероховатого ЭПР
Дифференцировка Т-лимфоцитов
Строение T-клеточного рецептора (TCR – “T-cell receptor”)
3 D модель T-клеточного рецептора
TCR – гетеродимеры, и или и цепей ( Т-клетки и Т-клетки) TCR тесно соединяется с CD 3 – комплексом полипептидных цепей, необходимых для трансдукции сигнала
Схематическая структура CD 4 (4 подобных Ig домена) и CD 8 (αβ – гетеродимер и αα- гомодимер)
В отличие от BCR, распознающего АГ в нативном виде, TCR связывает и распознает АГ только в составе молекул MHC. CD 4 и CD 8 классические корецепторы, распознающие комплекс «пептид-МНС» и участвующие в сигнальной трансдукции. Внеклеточные домены CD 4 и CD 8 связываются с консервативными МНС молекулами на АПК. CD 4 связывается с МНС класса II, а CD 8 – с МНС класса I
Распознавание антигена в составе молекул МНС I и II классов
Т-хелперы (Th) (CD 4+) Функциональная субпопуляция Т клеток, которые способствуют генерации цитотоксических Т лимфоцитов и участвуют в межклеточной кооперации с В-клетками, активируя их в направлении синтеза антител. Th распознают антиген в ассоциации с молекулами MHC класса II (феномен двойного распознавания)
Цитотоксические Т клетки (СD 8+) Клетки-киллеры, способные убивать инфицированные вирусом клетки, опухолевые клеткимишени, несущие на поверхности антигенные пептиды в комплексе с молекулами MHC класса I (феномен двойного распознавания, но для эффекторной функции)
Регуляторные Т-клетки (Treg) (CD 4+CD 25+Fox. P 3+) Т-клетки, зависимые от тимуса, регулирующие (супрессирующие) функцию других клеток иммунной системы
Основным процессом дифференцировки Т-лимфоцитов, также как и у В-лимфоцитов, является формирование многообразия антигенраспознающих Т-клеточных рецепторов
Локализация генов ТКР у человека
Реаранжировка (перестройка) генов V D J
Реаранжировка генов по принципу образования петли ДНК
Циркулярные образования ДНК в Т-клетках, недавно покинувших тимус
Этапы дифференцировки Т-лимфоцитов
Положительная и отрицательная селекция Т-лимфоцитов в тимусе
Положительная селекция тимоцитов
Отрицательная селекция тимоцитов
Положительная и отрицательная селекция тимоцитов
Спасибо за внимание.
Источник: https://present5.com/lekciya-4-differencirovka-t-i-v-limfocitov-2/
Виды Т-лимфоцитов: Т-киллеры, Т-хелперы, Т-супрессоры, Лимфоциты-амплификаторы, Т-клетки памяти
Эта группа клеток состоит из нескольких видов, также называемых субпопуляциями лимфоцитов. Долгое время выделяли только три их вида: это Т-лимфоциты хелперы, киллеры и супрессоры.
Однако в последние годы, начиная с 1990-2000-х, представление об их существующих разновидностях претерпело очередную трансформацию.
Помимо известных клеток, специалисты определили существование других типов: Т-клеток памяти и клеток-амплификаторов. Рассмотрим все существующие виды лимфоцитов подробнее.
Т-киллеры:
Т-киллеры, строго осуществляющие иммунный надзор, агрессивно реагируют на чужеродный белок. Именно они вызывают реакцию отторжения трансплантата при пересадке органов. По этой причине при пересадке человеку любого органа ему некоторое время дают специальные медикаменты, которые угнетают иммунную систему: уменьшают повышенное содержание лимфоцитов и нарушают их взаимодействие. Иначе любая подобная операция заканчивалась бы отторжением нового органа или ткани, а может, даже гибелью пациента, которому такое вмешательство проводится.
Интересен механизм работы этих клеток. В отличие от фагоцитов, активно атакующих, пожирающих и переваривающих чужеродные частицы, Т-киллеры ведут себя на первый взгляд достаточно сдержанно. Своими отростками они прикасаются к объекту, а затем разрывают контакт и «уходят по своим делам». Клетка же, к которой прикоснулся лимфоцит, спустя какое-то время погибает… Отчего?
Дело в том, что во время своего «смертельного поцелуя» Т-киллеры оставляют на поверхности уничтожаемой ими клетки частицы своей мембраны. В местах контакта частицы «разъедают» поверхность объекта нападения. В результате в обреченной на гибель клетке фактически образуется сквозное отверстие.
Она теряет ионы калия, внутрь нее входят ионы натрия и вода – так как клеточный барьер нарушается, ее внутренняя среда начинает напрямую сообщаться с внешней… В исходе клетка раздувается проникшей внутрь ее водой, из нее выходят белки цитоплазмы, органеллы разрушаются… Она погибает, а дальше к ней подходят фагоциты и пожирают ее остатки.
Вот такое страшное наказание готовит организм всем клеткам, которые были распознаны иммунитетом как «неправильные» или чужеродные.
Т-хелперы:
Задача хелперов на первый взгляд тоже довольно очевидна. Это клетки-помощники («help» значит «помогать»). А кому или чему они помогают? Они индуцируют, стимулируют иммунный ответ: под их влиянием усиливают свою работу цитотоксические лимфоциты. Также хелперы передают информацию о присутствии в теле чужеродного белка В-лимфоцитам, которые выделяют против них защитные антитела. Наконец, хелперы оказывают стимулирующее действие на работу фагоцитов, главным образом моноцитов.
 Это клетки-помощники («help» значит «помогать»). А кому или чему они помогают? Они индуцируют, стимулируют иммунный ответ: под их влиянием усиливают свою работу цитотоксические лимфоциты. Также хелперы передают информацию о присутствии в теле чужеродного белка В-лимфоцитам, которые выделяют против них защитные антитела. Наконец, хелперы оказывают стимулирующее действие на работу фагоцитов, главным образом моноцитов.
Это клетки-помощники («help» значит «помогать»). А кому или чему они помогают? Они индуцируют, стимулируют иммунный ответ: под их влиянием усиливают свою работу цитотоксические лимфоциты. Также хелперы передают информацию о присутствии в теле чужеродного белка В-лимфоцитам, которые выделяют против них защитные антитела. Наконец, хелперы оказывают стимулирующее действие на работу фагоцитов, главным образом моноцитов. - Т-хелперы
Т-супрессоры:
«Супрессия» означает «подавление». Если Т-лимфоциты хелперы усиливают реакцию иммунитета, то супрессоры, наоборот, подавляют. Притом эти клетки вовсе не устраивают саботаж иммунным процессам и не вредят нашему здоровью. Они просто регулируют силу иммунного ответа, что позволяет иммунной системе сдержанно и с умеренной силой отвечать на раздражители.
Лимфоциты-амплификаторы:
После того как в организм проник агрессор, в крови и тканях отмечается повышенное содержание лимфоцитов. Их число возрастает буквально в течение нескольких часов и может увеличиваться более чем в 2 раза. Отчего же нарастание количества клеток происходит так быстро? Просто в организме имеется их некоторый запас.
В селезенке и тимусе живут зрелые, полноценные лимфоциты. Их отличие от остальных состоит лишь в том, что они «не определились», к какому виду лимфоцитов принадлежат. Это и есть клетки-амплификаторы, при необходимости они участвуют в увеличении численности других Т-лимфоцитов.
Т-клетки памяти:
Справившись с очередной угрозой, лимфоциты ее запоминают. В организме человека образуется особый клон клеток, которые и хранят эти «воспоминания». Каждый клон несет в себе информацию об определенном виде угрозы. Если какой-то агрессор, с которым иммунная система уже встречалась, проникает в тело, то соответствующий клон размножается и быстро формирует вторичный иммунный ответ.
Разговор о разновидностях лимфоцитов и их функциях достаточно долгий. Здесь эта тема была представлена в максимально приемлемой и простой форме, без загрузки специфическими терминами и непонятными названиями. Будем надеяться, что любой читатель, даже не имеющий медицинского образования, примерно разобрался в том, как функционируют в его организме разные виды Т-лимфоцитов.
Из всего этого можно сделать очевидный вывод: для того чтобы жить полноценной здоровой жизнью, надо быть обладателем крепкой иммунной системы. Необходимо, чтобы процессы, о которых многие не задумываются, а еще большее количество людей даже не знает, происходили как надо.
Если природа не наградила вас устойчивым иммунитетом, вам следует задуматься о том, чтобы самостоятельно взяться за его укрепление. Для этого можно начать прием препарата Трансфер Фактор.
В нем содержатся информационные молекулы, при помощи которых лимфоциты в норме общаются друг с другом, управляют различными процессами и координируют их.
Восполняя недостаток естественных информационных молекул, средство является одним из самых рекомендуемых и эффективных препаратов для нормализации работы иммунитета, улучшения здоровья и профилактики заболеваний.
Клетки иммунитета обладают памятью и передают друг другу информацию
Источник: https://transferfaktory.ru/vidyi-t-limfotsitov
Дифференцировка t-лимфоцитов
В основе выделения этапов развития Т-лимфоцитов лежит состояние рецепторных V-генов и экспрессии TCR, а также корецепторов и других мембранных молекул. Схема дифференцировки Т-лимфоцитов (рис. 6-5) аналогична приведённой выше схеме развития В-лимфоцитов (см. рис. 5-13). Приведены ключевые характеристики фенотипа и ростовых факторов развивающихся Т-клеток.
Принятые обозначения стадий развития Т-клеток определяются экспрессией корецепторов: DN (от Double-Negative, CD4CD8) – двойные отрицательные, DP (от Double-Positive, CD4+CD8+) – двойные положительные, SP (от Single-Positive, CD4+CD8- и CD4CD8+) – одинарно положительные.
Деление DNтимоцитов на стадии DN1, DN2, DN3 и DN4 основывается на характере
Рис. 6-5. Развитие Т-лимфоцитов
экспрессии молекул CD44 и CD25. Другие условные обозначения: SCF (от Stem Cell Factor) – фактор стволовых клеток, lo (low; метка индекса) – низкий уровень экспрессии. Стадии реаранжировки: D-J – предварительный этап, соединение сегментов D и J (только в генах β- и δ-цепей TCR, см. рис. 6-2), V-DJ – завершающий этап, соединение зародышевого V-гена с объединённым сегментом DJ.
• Тимоциты дифференцируются из общей клетки-предшественника, которая ещё вне тимуса экспрессирует такие мембранные маркёры, как CD7, CD2, CD34 и цитоплазматическую форму CD3.
• Коммитированные к дифференцировке в T-лимфоциты клеткипредшественники мигрируют из костного мозга в субкапсулярную зону коры тимуса, где примерно в течение одной недели медленно пролиферируют. На тимоцитах появляются новые мембранные молекулы CD44 и CD25.
• Затем клетки перемещаются вглубь коры тимуса, молекулы CD44 и CD25 исчезают с их мембраны. В этой стадии начинается перестройка генов β-, γ- и δ-цепей TCR. Если гены γ- и δ-цепей успевают продуктивно, т.е.
без сдвига рамки считывания, перестроиться раньше, чем гены β-цепи, то лимфоцит дифференцируется далее как γδT.
В противном случае происходит экспрессия β-цепи на мембране в комплексе с pTα (инвариантной суррогатной цепью, заменяющей на этом этапе настоящую α-цепь) и CD3. Это служит
сигналом к прекращению перестройки генов γ- и δ-цепей. Клетки начинают пролиферировать и экспрессировать одновременно CD4 и CD8 – дважды позитивные тимоциты. При этом накапливается масса клеток с уже готовой β-цепью, но с ещё не перестроенными генами α-цепи, что вносит свой вклад в разнообразие αβ-гетеродимеров.
• На следующем этапе клетки перестают делиться и начинают перестраивать Vα-гены, причём несколько раз в течение 3-4 сут. Перестройка генов α-цепи приводит к необратимой делеции δ-локуса, расположенного между сегментами генов α-цепи.
• Происходят экспрессия TCR с каждым новым вариантом α-цепи и отбор (селекция) тимоцитов по силе связывания с комплексом MHC-пептид на мембранах эпителиальных клеток тимуса.
– Позитивная селекция: погибают тимоциты, не связавшие ни одного из доступных комплексов MHC-пептид. В результате позитивной селекции в тимусе погибает около 90\% тимоцитов.
– Негативная селекция уничтожает клоны тимоцитов, связывающих комплексы MHC-пептид со слишком высокой аффинностью. Негативная селекция элиминирует от 10 до 70\% клеток, прошедших позитивную селекцию.
– Тимоциты, связавшие какой-либо из комплексов MHC-пептид с правильной, т.е. средней по силе, аффинностью, получают сигнал к выживанию и продолжают дифференцировку.
• На короткое время с мембраны тимоцитов исчезают обе корецепторные молекулы, а затем экспрессируется одна из них: тимоциты, распознавшие пептид в комплексе с MHC-I, экспрессируют корецептор CD8, а с MHC-II – корецептор CD4. Соответственно на периферию выходят (в соотношении около 2:1) T-лимфоциты двух типов: CD8+ и CD4+, функции которых в предстоящих иммунных ответах различны.
– CD8+ T-клетки играют роль цитотоксических T-лимфоцитов (ЦТЛ) – они распознают и непосредственно убивают клетки, модифицированные вирусом, опухолевые и другие «изменённые» клетки (рис. 6-6).
– CD4+ T-клетки. Функциональная специализация CD4+ T-лимфоцитов более разнообразна. Значительная часть CD4+ T-лимфоцитов в процессе развития иммунного ответа становится T-хелперами (помощниками), взаимодействующими с В-лимфоцитами, Т-лимфоцитами и другими клетками при
Рис. 6-6. Механизм воздействия цитотоксического T-лимфоцита на клеткумишень.
В Т-киллере в ответ на увеличение концентрации Са2+ гранулы с перфорином (фиолетовые овалы) и гранзимами (жёлтые кружочки) сливаются с клеточной мембраной.
Освободившийся перфорин встраивается в мембрану клетки-мишени с последующим образованием пор, проницаемых для гранзимов, воды и ионов. В результате клетка-мишень лизируется
прямом контакте или через растворимые факторы (цитокины). В определённых случаях из них могут развиться CD4+ ЦТЛ: в частности, такие T-лимфоциты обнаружены в значительных количествах в коже больных с синдромом Лайелла.
Субпопуляции T-хелперов
С конца 80-х годов XX века было принято выделять 2 субпопуляции T-хелперов (в зависимости от того, какой набор цитокинов они продуцируют) – Th1 и Th2. В последние годы спектр субпопуляций CD4+ Т-клеток продолжает расширяться. Обнаружены такие субпопуляции, как: Th17, T-регуляторы, Tr1, Th3, Tfh и др.
Основные субпопуляции CD4+ Т-клеток:
• Th0 – CD4+ Т-лимфоциты на ранних стадиях развития иммунного ответа, они продуцируют только ИЛ-2 (митоген для всех лимфоцитов).
• Th1 – дифференцированная субпопуляция CD4+ Т-лимфоцитов, специализирующаяся на продукции ИФНγ, ФНОβ и ИЛ-2.
Эта субпопуляция осуществляет регуляцию многих реакций клеточного иммунитета, включая гиперчувствительность замедленного типа (ГЗТ) и активацию ЦТЛ.
Кроме того, Th1 стимулируют продукцию В-лимфоцитами опсонизирующих антител класса IgG, запускающих каскад активации комплемента. Развитие избыточного воспаления с последующим повреждением тканей напрямую связано с активностью Th1-субпопуляции.
• Th2 – дифференцированная субпопуляция CD4+ Т-лимфоцитов, специализирующаяся на выработке ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10 и ИЛ-13. Эта субпопуляция участвует в активации В-лимфоцитов и способствует секреции ими больших количеств антител разных классов, особенно IgE. Кроме того, Th2-субпопуляция участвует в активации эозинофилов и развитии аллергических реакций.
• Th17 – субпопуляция CD4+ Т-лимфоцитов, специализирующаяся на образовании ИЛ-17. Эти клетки осуществляют противогрибковую и антимикробную защиту эпителиальных и слизистых барьеров, а также играют ключевую роль в патологии аутоиммунных заболеваний.
• Т-регуляторы – CD4+ Т-лимфоциты, подавляющие активность других клеток иммунной системы посредством секреции иммуносупрессорных цитокинов – ИЛ-10 (ингибитора активности макрофагов и Th1-клеток) и ТФРβ – ингибитора пролиферации лимфоцитов.
Ингибиторный эффект может также достигаться при непосредственном межклеточном взаимодействии, поскольку на мембране некоторых Т-регуляторов экспрессированы индукторы апоптоза активированных и «отработавших» лимфоцитов – FasL (Fas-лиганд).
Существует несколько популяций CD4+ регуляторных Т-лимфоцитов: естественные (Treg), созревающие в тимусе (CD4+CD25+, экспрессируют фактор транскрипции Foxp3), и индуцированные – локализованные преимущественно в слизистых оболочках пищеварительного тракта и переключившиеся на образование ТФРβ (Th3) или ИЛ-10 (Tr1).
Нормальное функционирование Т-регуляторов необходимо для поддержания гомеостаза иммунной системы и предотвращения развития аутоиммунных заболеваний.
• Дополнительные хелперные популяции. В последнее время появляется описание всё новых популяций CD4+ Т-лимфоцитов, клас-
сифицированных по типу преимущественно продуцируемого ими цитокина. Так, как оказалось, одной из важнейших популяций являются Tfh (от англ. follicular helper – фолликулярный хелпер).
Эта популяция CD4+ Т-лимфоцитов преимущественно расположена в лимфоидных фолликулах и осуществляет хелперную функцию для В-лимфоцитов посредством продукции ИЛ-21, вызывая их созревание и терминальную дифференцировку в плазматические клетки.
Кроме ИЛ-21 Tfh могут также продуцировать ИЛ-6 и ИЛ-10, необходимые для дифференцировки В-лимфоцитов. Нарушение функций этой популяции приводит к развитию аутоиммунных заболеваний или иммунодефицитов. Другой «новоявленной» популяцией являются Th9 – продуценты ИЛ-9.
По-видимому, это Th2, переключившиеся на секрецию ИЛ-9, способного вызывать пролиферацию Т-хелперных клеток при отсутствии антигенной стимуляции, а также усиливать секрецию В-лимфоцитами IgM, IgG и IgE.
Основные субпопуляции Т-хелперов представлены на рис. 6-7. На рисунке суммированы современные представления об адаптивных субпопуляциях CD4+ Т-клеток, т.е. субпопуляций, формирующих-
Рис. 6-7. Адаптивные субпопуляции CD4+ Т-клеток (цитокины, дифференцировочные факторы, хемокиновые рецепторы)
ся при иммунном ответе, а не в ходе естественного развития клеток.
Для всех разновидностей Т-хелперов указаны цитокины-индукторы (на стрелках, ведущих к кружкам, символизирующим клетки), транскрипционные факторы (внутри кружков), хемокиновые рецепторы, направляющие миграцию (около линий, отходящих от «поверхности клетки»), и продуцируемые цитокины (в прямоугольниках, на которые направлены стрелки, отходящие от кружков).
Расширение семейства адаптивных субпопуляций CD4+ Т-клеток потребовало решения вопроса о природе клеток, с которыми взаимодействуют эти субпопуляции (кому они оказывают «помощь» в соответствии со своей функцией хелперов).
Эти представления отражены на рис. 6-8.
Здесь же представлен уточнённый взгляд на функции этих субпопуляций (участие в защите от определённых групп патогенов), а также о патологических последствиях несбалансированного усиления активности этих клеток.
Рис. 6-8. Адаптивные субпопуляции Т-клеток (клетки-партнёры, физиологические и патологические эффекты)
γδT-лимфоциты
Подавляющее большинство (99\%) T-лимфоцитов, проходящих лимфопоэз в тимусе, составляют αβT-клетки; менее 1\% – γδT-клетки. Последние в большинстве дифференцируются вне тимуса, в первую очередь в слизистых оболочках пищеварительного тракта.
В коже, лёгких, пищеварительном и репродуктивном трактах они являются доминирующей субпопуляцией внутриэпителиальных лимфоцитов. Среди всех T-лимфоцитов организма γδT-клетки составляют от 10 до 50\%.
В эмбриогенезе γδT-клетки появляются раньше αβT-клеток.
• γδT-клетки не экспрессируют CD4. Молекула CD8 экспрессирована на части γδT-клеток, но не в виде ap-гетеродимера, как на CD8+ apT-клетках, а в виде гомодимера из двух a-цепей.
• Антигенраспознающие свойства: γδTCR в большей степени напоминают иммуноглобулины, чем αβTCR, т.е. способны связывать нативные антигены независимо от классических молекул MHC – для γδT-клеток не обязателен или вовсе не нужен предварительный процессинг антигена АПК.
• Разнообразие γδTCR меньше, чем αβTCR или иммуноглобулинов, хотя в целом γδT-клетки способны распознавать широкий спектр антигенов (в основном это фосфолипидные антигены микобактерий, углеводы, белки теплового шока).
• Функции γδT-клеток ещё до конца не изучены, хотя становится преобладающим мнение, что они служат одним из связующих компонентов между врождённым и приобретённым иммунитетом. γδT-клетки – один из первых барьеров на пути патогенов. Кроме того, эти клетки, секретируя цитокины, играют важную иммунорегуляторную роль и способны дифференцироваться в ЦТЛ.
NKT-лимфоциты
Естественные киллерные Т-клетки (NKT-клетки) представляют особую субпопуляцию лимфоцитов, занимающую промежуточное положение между клетками врождённого и адаптивного иммунитета. Эти клетки имеют черты как NK-, так и Т-лимфоцитов.
NKT-клетки экспрессируют αβTCR и характерный для NK-клеток рецептор NK1.1, принадлежащий к суперсемье лектиновых гликопротеинов С-типа. Однако TCR-рецептор NKT-клеток имеет существенные отличия от TCR-рецептора обычных клеток.
У мышей большинство NKTклеток экспрессирует инвариантный V-домен a-цепи, состоящий из
сегментов Vα14-Jα18, иногда обозначаемый как Jα281. У человека V-домен α-цепи состоит из сегментов Vα24-JαQ. У мышей α-цепь инвариантного TCR преимущественно комплексируется с Vβ8.2, у человека – с Vβ11. Из-за особенностей строения цепей TCR NKTклеток называют инвариантным – iTCR.
Развитие NKT-клеток зависит от молекулы CD1d, которая имеет сходство с молекулами МНС-I. В отличие от классических молекул МНС-I, презентирующих Т-клеткам пептиды, CD1d презентирует Т-клеткам только гликолипиды.
Хотя считается, что печень является местом развития NKT-клеток, имеются строгие доказательства роли тимуса в их развитии. NKT-клетки играют важную роль в регуляции иммунитета. У мышей и людей с различными аутоиммунными процессами функциональная активность NKT-клеток сильно нарушена.
Полной картины значимости таких нарушений в патогенезе аутоиммунных процессов нет. При некоторых аутоиммунных процессах NKT-клетки могут играть супрессорную роль.
Помимо контроля аутоиммунных и аллергических реакций, NKTклетки участвуют в иммунном надзоре, вызывая при повышении функциональной активности отторжение опухолей.
Велика их роль в противомикробной защите, особенно на ранних этапах развития инфекционного процесса. NKT-клетки вовлекаются в различные воспалительные инфекционные процессы, особенно при вирусных поражениях печени.
В целом NKT-клетки – многофункциональная популяция лимфоцитов, несущая ещё много научных загадок.
На рис. 6-9 обобщены данные о дифференцировке Т-лимфоцитов на функциональные субпопуляции. Представлены несколько уровней бифуркации: γδТ/αβТ, далее для αβТ-клеток – NKT/ остальные Т-лимфоциты, для последних – CD4+/CD8+, для CD4+ Т-клеток – Th/Treg, для CD8+ Т-лимфоцитов – CD8αβ/CD8αα. Показаны также дифференцировочные транскрипционные факторы, ответственные за все линии развития.
Рис. 6-9. Естественные субпопуляции Т-лимфоцитов и их дифференцировочные факторы
Источник: https://yamedik.org/gematologiya/imyn_str_fi_hi/dyfferencyrovka_lymfocytov/
Развитие, дифференцировка
Важную роль в их развитии играет тимус. Дифференцировка Т-лимфоцитов начинается уже в период эмбрионального развития. В вилочковой железе создается особое микроокружение за счет тимического эпителия, влияющее на дифференцировку Т-лимфоцитов.
Предшественники Т-лимфоцитов заселяют вилочковую железу, пролиферируют там, превращаются в Т-лимфоциты, а затем мигрируют в тимусзависимые зоны периферические органы иммунной системы, где приобретают способность к специфической активации. Под влиянием стромальных элементов лимфоциты приобретают способность реагировать на стимулы микроокружения, т.е. на тимические факторы, и синтезировать специфические маркеры тимоцитов и Т-лимфоцитов.
Процесс пролиферации лимфоцитов в вилочковой железе протекает очень интенсивно, но не все клетки мигрируют из вилочковой железы в виде Т-лимфоцитов: большинство из них там же погибает. Полагают, что причиной их гибели служит присоединение антигена к антигенспецифическому рецептору этих клеток.
В вилочковой железе нет чужеродных антигенов, вот почему этот механизм может служить для удаления Т-лимфоцитов, способных реагировать с аутоструктурами организма, т.е. защищать его от аутоиммунных реакций.
Специфическое взаимодействие с антигеном служит началом процесса дифференцировки в зрелые и долгоживущие клетки, составляющие большую часть рециркулирующих Т-лимфоцитов.
Т-лимфоцит, аналогично В-лимфоциту, несет на своей поверхности специфические рецепторы для распознавания антигенов. Структурная основа процесса узнавания антигена до сих пор неизвестна.[13]
У всех иммунокомпетентных Т-лимфоцитов антигенный рецептор нековалентно, но достаточно прочно связан в комплекс с молекулой Т3, состоящей из пяти пептидных цепей, которая участвует в передаче сигнала от узнающего антиген гетеродимера внутрь клетки. Таким образом, рецептор Т-лимфоцитов представляет собой комплекс, образованный гетеродимером и Т3 (или СД3). [16]
Субпопуляции Т-лимфоцитов, их функции
Периферические Т-клетки подразделяются на различные субпопуляции, значительный прогресс изучения которых произошел с помощью техники моноклональных антител.
Т-хелперы (помощники, индукторы) — запрограммированы индуцировать размножение и (или) дифференцировку клеток других типов. Например, они индуцируют секрецию антител В-лимфоцитами и стимулируют моноциты, тучные клетки и предшественники Т-киллеров к участию в клеточных иммунных реакциях.
Т-хелперы активируют эффекторные, а Т-индукторы — регуляторные клетки, хотя различие между ними оказывается искусственным.
Эта субпопуляция активируется антигенами, ассоциируемыми с продуктами генов МНС класса II — молекулами класса II, представленными преимущественно на поверхности В-клеток и макрофагов.
Существует как минимум три класса Т-хелперов:
1. Т-хелперы, узнающие МНС. Они обладают специфичностью к антигену, презентированному в комплексе со своими молекулами МНС класса II.
Эти клетки пролиферируют в ответ на комплекс «антиген+молекулы МНС класса II», а также индуцируют пролиферацию В-клеток, связавших данный антиген и имеющих на своей поверхности молекулы МНС II класса, а также дифференцировку этих В-клеток до антителообразующих клеток;
2. Т-хелперы, узнающие иммуноглобулины. Некоторые Т-хелперы обладают специфичностью как к антигенам, так и к собственным идиотипическим детерминантам. Они активируют В-клетки, имеющие такие же идиотипические детерминанты.
Эти же Т-клетки узнают собственные идиотипы в комплексе со своими молекулами МНС класса II и, возможно, могут активировать В-клетки в отсутствие антигена и других Т-клеток.
Физиологическая роль этих клеток точно неизвестна;
3. Т-хелперы, секретирующие лимфокины. Часть Т-клеток осуществляет свою «хелперную» функцию через освобождение «дальнодействующих» факторов — ИЛ-2 и фактора, замещающего Т-клетки. Эти факторы называются лимфокинами.
Они активируют или способствуют активации других Т- или В-клеток, узнавая антиген в комплексе с собственными белками МНС класса II. Эти же клетки могут индуцировать реакции гиперчувствительности замедленного типа.
[14]
Источник: https://studbooks.net/828214/meditsina/razvitie_differentsirovka
Дифференцировка Т-лимфоцитов
Стволовая гемопоэтическая клетка, мигрирующая в тимус, превращается (дифференцируется) под влиянием тимического микроокружения в Т-лимфоцит. Цель дифференцировки:
- обучить распознаванию чужеродного материала, попавшего в организм, и его разрушению (т. е. осуществлению киллинг-эффекта);
- создать толерантность по отношению к собственным (self) антигенам. тимус играет главную роль в этих процессах, поскольку является тем органом, где про¬исходит антигеннезависимая дифференцировка Т-клеток и создание (генерирование) чрезвычайно разнообразного набора (репертуара) антигенраспознающих Т-клеточных рецепторов.
Вначале стволовая гемопоэтическая клетка попадает в корковую зону тимуса и превращается в ранний предшественник Т-лимфоцита. Фенотип этой клетки следующий: ТАГРР-альфа, бета +, CD3+ CD4-, CD8-, т. е. характеризуется наличием Т-клеточного распознающего рецептора, в составе которого имеются альфа- и бета-цепи, CD3 структура, но отсутствуют молекулы CD4 и CD8.
Далее, здесь же в корковой зоне тимуса, под влиянием тимического микроокружения, гормонов тимуса и, особенно, ИЛ-7 ранний предшественник Т-лимфоцита превращается в незрелый Т-лимфоцит, фенотип которого следующий: ТАГРР-альфа, бета +, CD3+, CD4+, CD8+.
Набор таких мембранных структур говорит о том, что данная клетка способна: 1) распознать любой антиген с помощью ТАГРР-альфа, бета; 2) после распознавания передать сигнал внутрь клетки для ее активации с помощью CD3 структуры: 3) превратиться как в CD4+ (хелпер), так и в CD8+ (киллер) клетки при развитии эффекторного звена иммунного ответа.
На следующем этапе дифференцировки незрелый предшественник Т-лимфоцита переходит в мозговое вещество тимуса, где завершается тимический этап созревания.
При этом происходят два важнейших события: 1) индуцируется толерантность к аутоантигенам; таким образом, минимизируется возможность развития аутоиммунного заболевания; 2) происходит разделение Т-лимфоцитов на две субпопуляции: CD4+CD8- (хелперы) и CD4-CD8+ (киллеры) (не нужно за-бывать, что на их мембране сохраняются молекулы ТАГРР-альфа, бета и CD3). Этот этап также реализуется при важном участии ИЛ-7.
Покидая тимус, зрелые покоящиеся Т-лимфоциты, которые находятся в G(O) стадии клеточного цикла, расселяются в Т-зоны периферических лимфоидных органов. Такие Т-лимфоциты характеризуется следующими свойствами:
- способностью распознавать чужеродные антигены, которые презентируются ему в виде пептида с помощью молекул ГКГ класса I и класса II, и развивать эфферентную часть иммунного ответа;
- неспособностью распознавать большинство аутологичных антигенов, как в растворимой форме, так и в виде молекул на мембране клеток. Это главное препятствие на пути к развитию аутоиммунного ответа.
Часть Т-лимфоцитов, покидающих тимус, все же способна распознавать аутоантигены, однако такие Т-лимфоциты (и В-лимфоциты) либо подвергаются делении (разрушению) в периферических органах, либо находятся в состоянии анергии (неспособности к активации и реализации эфферентной части иммунного ответа).
Т-лимфоциты-хелперы (CD4+ клетки) представлены тремя субпопуляциями: т. н. нулевыми Т-хелперами (Тх0), которые дифференцируются в Т-хелперы 1-го типа (Tx1) и 2-го типа (Тх2). В этой дифференцировке основную роль играют ИЛ-12, ИЛ-2, гамма-интерферон, ИЛ-10, ИЛ-4, ИЛ-5.
Т-лимфоциты-хелперы (CD4+ клетки) участвует в распознавании атигенногопептида , который презентируется с помощью молекул ГКГ класса П. В этом случае для активации Т-лимфоцита необходим дополнительный, костимуляционный, сигнал.
Источник: https://www.eurolab.ua/encyclopedia/immunology-and-allergy/47671/
Иммунная система:
Основными клетками, осуществляющими иммунные реакции, являются Т- и В-лимфоциты (и производные последних – плазмоциты), макрофаги, а также ряд взаимодействующих с ними клеток (тучные клетки, эозинофилы и др.).
Лимфоциты
Популяция лимфоцитов функционально неоднородна. Различают три основных вида лимфоцитов: Т-лимфоциты, В-лимфоциты и так называемые нулевые лимфоциты (0-клетки).
Лимфоциты развиваются из недифференцированных лимфоидных костномозговых предшественников и при дифференцировке получают функциональные и морфологические признаки (наличие маркеров, поверхностных рецепторов), выявляемые иммунологическими методами.
0-лимфоциты (нулевые) лишены поверхностных маркеров и рассматриваются как резервная популяция недифференцированных лимфоцитов.
Т-лимфоциты — самая многочисленная популяция лимфоцитов, составляющая 70—90% лимфоцитов крови.
Они дифференцируются в вилочковой железе — тимусе (отсюда их название), поступают в кровь и лимфу и заселяют Т-зоны в периферических органах иммунной системы — лимфатических узлах (глубокая часть коркового вещества), селезенке (периартериальные влагалища лимфоидных узелков), в одиночных и множественных фолликулах различных органов, в которых под влиянием антигенов образуются Т-иммуноциты (эффекторные) и Т-клетки памяти. Для Т-лимфоцитов характерно наличие на плазмолемме особых рецепторов, способных специфически распознавать и связывать антигены. Эти рецепторы являются продуктами генов иммунного ответа [Ir-гены, (immune response)]. Т-лимфоциты обеспечивают клеточный иммунитет, участвуют в регуляции гуморального иммунитета, осуществляют продукцию цитокинов при действии антигенов.
В популяции Т-лимфоцитов различают несколько функциональных групп клеток: цитотоксические лимфоциты (Тц), или Т-киллеры (Тк), Т-хелперы (Тх), Т-супрессоры (Тс).
Тк участвуют в реакциях клеточного иммунитета, обеспечивая разрушение (лизис) чужеродных клеток и собственных измененных клеток (например, опухолевых клеток). Рецепторы позволяют им распознавать белки вирусов и опухолевых клеток на их поверхности.
При этом активизация Тц (киллеров) происходит под влиянием антигенов гистосовместимости на поверхности чужеродных клеток.
Кроме того, Т-лимфоциты участвуют в регуляции гуморального иммунитета с помощью Тх и Тс. Тх стимулируют дифференцировку В-лимфоцитов, образование из них плазмоцитов и продукцию иммуноглобулинов (Ig).
Tx имеют поверхностные рецепторы, которые связываются с белками на плазмолемме В-клеток и макрофагов, стимулируя Тх и макрофаги к пролиферации, продукции интерлейкинов (пептидных гормонов), а В-клетки — к продукции антител.
Таким образом, главной функцией Тх является распознавание чужеродных антигенов (представляемых макрофагами), секреция интерлейкинов, стимулирующих В-лимфоциты и другие клетки для участия в иммунных реакциях.
Снижение в крови числа Тх ведет к ослаблению защитных реакций организма (эти лица более подвержены инфекциям). Отмечено резкое снижение числа Тх у лиц, инфицированных вирусом СПИДа.
Тс способны ингибировать активность Тх, В-лимфоцитов и плазмоцитов. Они участвуют в аллергических реакциях, реакциях гиперчувствительности. Тс подавляют дифференцировку В-лимфоцитов.
Одной из основных функций Т-лимфоцитов является продукция цитокинов, которые оказывают стимулирующее или тормозящее влияние на клетки, участвующие в иммунном ответе (хемотаксические факторы, макрофаги ингибирующий фактор — МИФ, неспецифические цитотоксические вещества и др.).
Натуральные киллеры. Среди лимфоцитов в крови, кроме вышеописанных Тц, выполняющих функцию киллеров, имеются так называемые натуральные киллеры (Нк, NK), которые также участвуют в клеточном иммунитете. Они образуют первую линию защиты против чужеродных клеток, действуют немедленно, быстро разрушая клетки.
Нк в собственном организме разрушают опухолевые клетки и клетки, инфицированные вирусом. Тц образуют вторую линию защиты, так как для их развития из неактивных Т-лимфоцитов требуется время, поэтому они вступают в действие позже Нк.
Нк — это большие лимфоциты диаметром 12—15 мкм, имеют дольчатое ядро и азурофильные гранулы (лизосомы) в цитоплазме.
В-лимфоциты являются основными клетками, участвующими в гуморальном иммунитете. У человека они образуются из СКК красного костного мозга, затем поступают в кровь и далее заселяют В-зоны периферических лимфоидных органов — селезенки, лимфатических узлов, лимфоидные фолликулы многих внутренних органов. В крови их содержится 10—30% от всей популяции лимфоцитов.
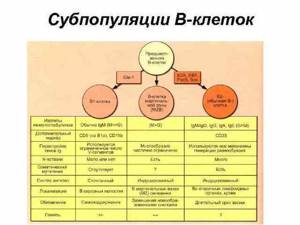
Для В-лимфоцитов характерно наличие на плазмолемме поверхностных иммуноглобулиновых рецепторов (SIg или MIg) для антигенов. Каждая В-клетка содержит 50000…150000 антигенспецифических молекул SIg.
В популяции В-лимфоцитов находятся клетки с различными SIg: большинство (⅔) содержат IgM, меньшее число (⅓) — IgG и около 1—5 % — IgA, IgD, IgE.
В плазмолемме В-лимфоцитов имеются также рецепторы для комплемента (С3) и Fc-рецепторы.
При действии антигена В-лимфоциты в периферических лимфоидных органах активизируются, пролиферируют, дифференцируются в плазмоциты, активно синтезирующие антитела различных классов, которые поступают в кровь, лимфу и тканевую жидкость.
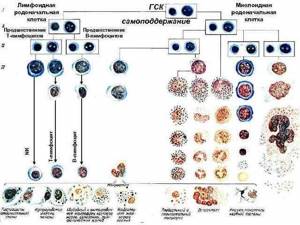
Развитие Т- и В-лимфоцитов
Родоначальником всех клеток иммунной системы является кроветворная стволовая клетка (СКК). СКК локализуются в эмбриональном периоде в желточном мешке, печени, селезенке.
В более поздний период эмбриогенеза они появляются в костном мозге и продолжают пролиферировать в постнатальной жизни.
Из СКК в костном мозге образуется клетка—предшественник лимфопоэза (лимфоидная мультипотентная родоначальная клетка), которая генерирует два типа клеток: пре-Т-клетки (предшественники Т-клеток) и пре-В-клетки (предшественники В-клеток).
Дифференцировка Т-лимфоцитов
Пре-Т-клетки мигрируют из костного мозга через кровь в центральный орган иммунной системы — вилочковую железу (тимус). Еще в период эмбрионального развития в вилочковой железе создается микроокружение, имеющее значение для дифференцировки Т-лимфоцитов.
В формировании микроокружения особая роль отводится ретикулоэпителиальным клеткам этой железы, способным к продукции ряда биологически активных веществ. Мигрирующие в вилочковую железу пре-Т-клетки приобретают способность реагировать на стимулы микроокружения.
Пре-Т-клетки в вилочковой железе пролиферируют, трансформируются в Т-лимфоциты, несущие характерные мембранные антигены (CD4+, CD8+). Т-лимфоциты генерируют и «поставляют» в кровообращение и в тимусзависимые зоны периферических лимфоидных органов 3 типа лимфоцитов: Тц, Тх и Тс. Мигрирующие из вилочковой железы «девственные» Т-лимфоциты (виргильные Т-лимфоциты) являются короткоживущими.
Специфическое взаимодействие с антигеном в периферических лимфоидных органах служит началом процессов их пролиферации и дифференцировки в зрелые и долгоживущие клетки (Т-эффекторные и Т-клетки памяти), составляющие большую часть рециркулирующих Т-лимфоцитов.
Из вилочковой железы мигрируют не все клетки. Часть Т-лимфоцитов погибает. Существует мнение, что причиной их гибели служит присоединение антигена к антигенспецифическому рецептору.
В вилочковой железе нет чужеродных антигенов, поэтому данный механизм может служить для удаления Т-лимфоцитов, способных реагировать с собственными структурами организма, т.е. выполнять функцию защиты от аутоиммунных реакций.
Гибель части лимфоцитов является генетически запрограммированной (апоптоз).
Дифференцировочные антигены Т-клеток. В процессе дифференцировки лимфоцитов на их поверхности появляются специфические мембранные молекулы гликопротеидов. Такие молекулы (антигены) можно обнаружить с помощью специфических моноклональных антител.
Получены моноклональные антитела, которые реагируют лишь с одним антигеном клеточной мембраны. С помощью набора моноклональных антител можно идентифицировать субпопуляции лимфоцитов. Имеются наборы антител к дифференцировочным антигенам лимфоцитов человека.
Антитела составляют относительно немного групп (или «кластеров»), каждая из которых узнает один единственный белок клеточной поверхности. Создана номенклатура дифференцировочных антигенов лейкоцитов человека, выявляемых моноклональными антителами.
Эта CD-номенклатура (CD — cluster of differentiation — кластер дифференцировки) базируется на группах моноклональных антител, реагирующих с одними и теми же дифференцировочными антигенами.
Получены многоклональные антитела к ряду дифференцировочных антигенов Т-лимфоцитов человека. При определении общей популяции Т-клеток могут быть использованы моноклональные антитела специфичностей CD (CD2, CD3, CDS, CD6, CD7).
Известны дифференцировочные антигены Т-клеток, которые характерны либо для определенных стадий онтогенеза, либо для различающихся по функциональной активности субпопуляций. Так, CD1 — маркер ранней фазы созревания Т-клеток в вилочковой железе.
В процессе дифференцировки тимоцитов на их поверхности экспрессируются одновременно маркеры CD4 и CD8. Однако в последующем маркер CD4 исчезает с части клеток и сохраняется только на субпопуляции, переставшей экспрессировать антиген CD8. Зрелые CD4+ клетки являются Тх.
Антиген CD8 экспрессируется примерно на ⅓ периферических Т-клеток, которые созревают из CD4+/CD8+ Т-лимфоцитов. Субпопуляция CD8+ Т-клеток включает цитотоксические и супрессорные Т-лимфоциты.
Антитела к гликопротеинам CD4 и CD8 широко используются для того, чтобы различать и разделять Т-клетки соответственно на Тх и Тц.
Кроме дифференцировочных антигенов, известны специфические маркеры Т-лимфоцитов.
Т-клеточные рецепторы для антигенов представляют собой антителоподобные гетеродимеры, состоящие из полипептидных α- и β-цепей.
Каждая из цепей имеет длину в 280 аминокислот, большая внеклеточная часть каждой цепи свернута в два Ig-подобных домена: один вариабельный (V) и один константный (С).
Антителоподобный гетеродимер кодируется генами, которые собираются из нескольких генных сегментов в процессе развития Т-клеток в вилочковой железе.
Дифференцировка В-лимфоцитов
Предшественники В-клеток (пре-В-клетки) развиваются в дальнейшем у птиц в фабрициевой сумке (bursa), откуда произошло название В-лимфоциты, у человека и млекопитающих — в костном мозге.
Сумка Фабрициуса (bursa Fabricii) — центральный орган иммунопоэза у птиц, где происходит развитие В-лимфоцитов, находится в области клоаки. Для ее микроскопического строения характерно наличие многочисленных складок, покрытых эпителием, в которых расположены лимфоидные узелки, ограниченные мембраной.
В узелках содержатся эпителиоциты и лимфоциты на различных стадиях дифференцировки. В период эмбриогенеза в центре фолликула формируется мозговая зона, а на периферии (снаружи от мембраны) — корковая зона, в которую, вероятно, мигрируют лимфоциты из мозговой зоны.
В связи с тем что в сумке Фабрициуса у птиц образуются исключительно В-лимфоциты, она является удобным объектом для изучения строения и иммунологических характеристик этого вида лимфоцитов. Для ультрамикроскопического строения В-лимфоцитов характерно наличие в цитоплазме групп рибосом в виде розеток.
Эти клетки имеют более крупные ядра и менее плотный хроматин, чем у Т-лимфоцитов, в связи с увеличением содержания эухроматина.
В-лимфоциты отличаются от других типов клеток способностью синтезировать иммуноглобулины. Зрелые В-лимфоциты экспрессируют Ig на клеточной мембране. Такие мембранные иммуноглобулины (MIg) функционируют как антигенспецифические рецепторы.
Пре-В-клетки синтезируют внутриклеточный цитоплазматический IgM, но не имеют поверхностных иммуноглобулиновых рецепторов. Костномозговые виргильные В-лимфоциты имеют IgM-рецепторы на своей поверхности. Зрелые В-лимфоциты несут на своей поверхности иммуноглобулиновые рецепторы различных классов — IgM, IgG и др.
Дифференцированные В-лимфоциты поступают в периферические лимфоидные органы, где при действии антигенов происходят пролиферация и дальнейшая специализация В-лимфоцитов с образованием плазмоцитов и В-клеток памяти (ВП).
В ходе своего развития многие В-клетки переключаются с выработки антител одного класса на выработку антител других классов. Этот процесс называется переключением класса.
Все В-клетки начинают свою деятельность по синтезу антител с выработки молекул IgM, которые встраиваются в плазматическую мембрану и служат рецепторами для антигена. Затем, еще до взаимодействия с антигеном, большая часть В-клеток переходит к одновременному синтезу молекул IgM и IgD.
Когда виргильная В-клетка переходит от выработки одного лишь мембраносвязанного IgM к одновременному синтезу мембраносвязанных IgM и IgD, переключение происходит, вероятно, благодаря изменению процессинга РНК.
При стимуляции антигеном некоторые из этих клеток активируются и начинают выделять антитела IgM, преобладающие в первичном гуморальном ответе.
Другие стимулированные антигеном клетки переключаются на выработку антител классов IgG, IgE или IgA; В-клетки памяти несут эти антитела на своей поверхности, а активные В-клетки их секретируют. Молекулы IgG, IgE и IgA в совокупности называются антителами вторичных классов, так как они, по-видимому, образуются только после антигенной стимуляции и преобладают во вторичных гуморальных ответах.
При помощи моноклональных антител удалось выявить определенные дифференцировочные антигены, которые еще до появления цитоплазматических µ-цепей позволяют отнести несущий их лимфоцит к В-клеточной линии. Так, антиген CD19 является самым ранним маркером, позволяющим отнести лимфоцит к В-клеточному ряду. Он присутствует на пре-В-клетках в костном мозге, на всех периферических В-клетках.
Антиген, выявляемый моноклональными антителами группы CD20, специфичен для В-лимфоцитов и характеризует более поздние стадии дифференцировки.
На гистологических срезах антиген CD20 выявляется на В-клетках герминативных центров лимфоидных узелков, в корковом веществе лимфатических узлов. В-лимфоциты несут также ряд других (например, CD24, CD37) маркеров.
Различают антигеннезависимую и антигензависимую дифференцировку и специализацию В- и Т-лимфоцитов.
Антигеннезависимая пролиферация и дифференцировка генетически запрограммированы на образование клеток, способных давать специфический тип иммунного ответа при встрече с конкретным антигеном благодаря появлению на плазмолемме лимфоцитов особых «рецепторов». Она совершается в центральных органах иммунитета (тимус, костный мозг или фабрициева сумка у птиц) под влиянием специфических факторов, вырабатываемых клетками, формирующими микроокружение (ретикулярная строма или ретикулоэпителиальные клетки в тимусе).
Антигензависимая пролиферация и дифференцировка Т- и В-лимфоцитов происходят при встрече с антигенами в периферических лимфоидных органах, при этом образуются эффекторные клетки и клетки памяти (сохраняющие информацию о действовавшем антигене).
Образующиеся Т-лимфоциты составляют пул долгоживущих, рециркулирующих лимфоцитов, а В-лимфоциты — короткоживущих клеток.
Некоторые термины из практической медицины:
- вакцина (vaccinum; лат. vaccinus коровий) — препарат, получаемый из живых аттенуированных штаммов или убитых культур микроорганизмов, их токсинов или антигенов, предназначенный для активной иммунизации людей и животных;
- анавакцина (anavaccinum; ана- + вакцина) — убитая вакцина, приготовленная путем продолжительного воздействия на взвесь микробов раствора формалина;
- вакцинация (vaccinatio; франц. vaccination) — метод создания активного иммунитета против инфекционной болезни путем введения вакцины в организм человека или животного;
- иммунизация (1) (immunisatio; лат. immunis свободный, избавленный от чего-либо; син.: прививки) — противоэпидемическое мероприятие, основанное на регламентированном по времени и возрастным группам применении средств специфической профилактики инфекционных болезней;
- иммунизация (2) (immunisatio) введение вакцин (анатоксинов) животным-продуцентам и людям — донорам крови с целью получения от них иммунных сывороток (иммуноглобулинов);
- иммунитет активный иммунитет, возникающий в результате иммунного ответа организма на введение антигена;
- иммунитет пассивный иммунитет, возникающий при передаче антител в форме антисыворотки или иммуноглобулинов от иммунизированного донора, а также от матери через плаценту или с молозивом;
- иммунологическая недостаточность (син.: иммунодефицитное состояние, иммунопарез, иммунодефицит) — снижение или отсутствие иммунного ответа вследствие дефекта системы иммунитета;
Источник: https://morphology.dp.ua/_mp3/immunity2.php